Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional
https://doi.org/10.19137/cienvet202426104
Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional
ARTICULO DE INVESTIGACION
Effect of alpha-lipoic acid on rat brain and heart microsomes subjected to oxidative stress induced by ascorbate-Fe++
Efecto del ácido alfa-lipoico en microsomas de cerebro y corazón de rata sometidos a estrés oxidativo inducido por ascorbato-Fe++
Efeito do ácido alfa-lipóico em microssomas cerebrais e cardíacos de ratos submetidos ao estresse oxidativo induzido por ascorbato-Fe++
Gavazza MB1, Marmunti ME1, Palacios A1
1Department of Biochemistry, Faculty of Veterinary Sciences, National University of La Plata, CC 296, 1900, La Plata, Buenos Aires, Argentina.
Fecha de recepción: 18 de agosto de 2023 /Fecha de aceptado para su publicación: 22 de noviembre de 2023
Abstract: Oxidative stress is caused by the overproduction of reactive oxygen species that generate an imbalance in the cellular antioxidant capacity. In this study, the effect of alpha-lipoic acid on microsomes isolated from rat brain and heart was analyzed by determining chemiluminescence (expressed as counts per minute) and thiobarbituric acid reactive substances. Oxidative stress was induced by subjecting the samples to an ascorbate-Fe++-dependent pro-oxidant system at 37 °C for 120 minutes. The inhibitory effect of alpha-lipoic acid was compared using different concentrations thereof, corresponding to 50, 150 and 250 µg per mg of microsomal protein. Controls were performed simultaneously without the addition of the pro-oxidant. A significant increase in light emission and malondialdehyde was observed in the microsomes of both organs in the ascorbate-Fe++ group. Analysis of chemiluminescence levels and thiobarbituric acid reactive substances indicated that alpha-lipoic acid acted as an antioxidant that protected rat heart microsomes from damage by lipid peroxidation at all doses tested. In brain microsomes, alpha-lipoic acid was observed to act as an antioxidant only at the 150 µg/ml dose. In the latter case, it will be necessary to test new doses of it to demonstrate the effects on these membranes. In conclusion, alpha-lipoic acid acted as an antioxidant to protect the membranes of both organs against peroxidative damage.
Keywords: Alpha-lipoic acid, Oxidative stress, Microsomes, Brain, Heart
Resumen: El estrés oxidativo es causado por la sobreproducción de especies reactivas de oxígeno que generan un desequilibrio en la capacidad antioxidante celular. En este estudio, se analizó mediante la determinación de la quimioluminiscencia (expresada como cuentas por minuto) y sustancias reactivas del ácido tiobarbitúrico, el efecto del ácido alfa-lipoico en microsomas aislados de cerebro y corazón de rata. El estrés oxidativo se indujo sometiendo a las muestras a un sistema prooxidante ascorbato-Fe++-dependiente a 37 °C durante 120 minutos. Se comparó el efecto inhibitorio del ácido alfa-lipoico utilizando diferentes concentraciones del mismo, correspondientes a 50, 150 y 250 µg por mg de proteína microsomal. Se realizaron simultáneamente controles sin la adición del prooxidante. Se observó en los microsomas de ambos órganos un aumento significativo de la emisión lumínica y el malondialdehído en el grupo ascorbato-Fe++. El análisis en los niveles de quimioluminiscencia y sustancias reactivas del ácido tiobarbitúrico indicó que el ácido alfa-lipoico actuó como un antioxidante que protegió a los microsomas del corazón de rata del daño por la peroxidación lipídica en todas las dosis ensayadas. En microsomas cerebrales, se observó que el ácido alfa-lipoico actuó como antioxidante solo a la dosis de 150 µg/ml. En este último caso, será necesario probar nuevas dosis del mismo para demostrar los efectos sobre estas membranas. En conclusión, el ácido alfa-lipoico actuó como un antioxidante protector de las membranas de ambos órganos frente al daño peroxidativo.
Palabras claves: Ácido alfa lipoico; Estrés Oxidativo; Microsomas; Cerebro; Corazón
Resumo. O estresse oxidativo é causado pela superprodução de espécies reativas de oxigênio que geram um desequilíbrio na capacidade antioxidante celular. Neste estudo, o efeito do ácido alfa-lipóico em microssomas isolados de cérebro e coração de ratos foi analisado pela determinação da quimioluminescência (expressa em contagens por minuto) e substâncias reativas ao ácido tiobarbitúrico. O estresse oxidativo foi induzido submetendo as amostras a um sistema pró-oxidante dependente de ascorbato-Fe++ a 37 °C por 120 minutos. O efeito inibitório do ácido alfa-lipóico foi comparado usando diferentes concentrações do mesmo, correspondendo a 50; 150 e 250 µg por mg de proteína microssomal. Os controles foram realizados simultaneamente sem a adição do pró-oxidante. Um aumento significativo na emissão de luz e malondialdeído foi observado nos microssomos de ambos os órgãos no grupo ascorbato-Fe++. A análise dos níveis de quimioluminescência e das substâncias reativas ao ácido tiobarbitúrico indicou que o ácido alfa-lipóico atuou como um antioxidante que protegeu os microssomos cardíacos de ratos contra danos por peroxidação lipídica em todas as doses testadas. Nos microssomas cerebrais, observou-se que o ácido alfa-lipóico atua como antioxidante apenas na dose de 150 µg/ml. Neste último caso, será necessário testar novas doses para demonstrar os efeitos nessas membranas. Em conclusão, o ácido alfa-lipóico atuou como um antioxidante para proteger as membranas de ambos os órgãos contra danos peroxidativos.
Palavras-chave: Ácido alfa-lipóico; Estresse oxidativo; microssomas; Cérebro; Coração
Introduction
Oxidative stress is considered to be the imbalance between the formation of reactive oxygen species (ROS) and cellular antioxidant capacity that is the result of increased ROS generation and/or dysfunction of the antioxidant system(1). ROS can be generated from several cellular sources including nicotinamide adenine dinucleotide phosphate oxidase (NOX), xanthine oxidase, and the mitochondrial electron transport chain(2). It is important to note that ROS production is a normal physiological process and is essential for redox balance; however, uncontrolled overproduction of ROS has been found to induce protein and lipid peroxidation as well as DNA mutagenesis that can lead to irreversible cell damage or death(3). The brain is vital part of the organism functioning as coordinating and regulating-system for body parts. It is considered highly vulnerable to oxidative stress than other organs of the body as it consumes high amount of oxygen, contains high amounts of polyunsaturated fatty acid and low levels of antioxidant enzymes(4).
Antioxidant compounds are one of the systems that the body has to counteract reactive species and free radicals(5). When present in low concentrations, antioxidants significantly inhibit the oxidative process of ROS by oxidizing themselves, and stop radical chain reactions by eliminating intermediates. Therefore, they are substances capable of protecting cells from oxidative stress by inhibiting or slowing down the effect of free radicals by reacting with them, to form a more stable radical that does not tend to react with oxygen(6). Alpha-lipoic acid (ALA) is present in both plants and animals. ALA contains 8 carbons with 2 sulfur atoms in dithiol ring. In the liver, ALA can be synthesized naturally via several enzymatic reactions from octanoic acid and cysteine(7). ALA is absorbed via various carriers, including monocarboxylic and sodium-dependent multivitamin carriers. The oxidized and reduced forms of dihydrolipoic acid (DHLA), both of which are antioxidants and anti-inflammatory agents, are responsible for the effects of ALA on the cellular mechanism(8, 9). Recently, ALA attracted enormous attention in neurodegenerative disease management as a potential therapeutic option(10).
The present study was designed to determine whether rat brain and heart microsomes could be targets for non-enzymatic lipid peroxidation, as well as to establish the level of protection of such membranes incubated with ALA. For this, oxidative stress was induced in both microsomes by subjecting samples (1 mg of protein) to a pro-oxidant system dependent on ascorbate-Fe++-(inducer) (0.4 mM)-Fe++ (2.15 μM) at 37 °C for 120 minutes. The brain was selected for its high capacity to produce free radicals and the heart for its high partial pressure of oxygen. In aqueous media, ascorbic acid is an antioxidant at concentration levels of the order of 1 mM and acts as a pro-oxidant at low concentrations (0.4 mM), especially in the presence of metal ions(11). The degradation process was followed by thiobarbituric acid reactive substances (TBARS) and by chemiluminescence (CL) determination(12). Measurement of light emission from a chemical reaction is very useful from an analytical point of view because, under appropriate experimental conditions, light output is directly related to analytical concentration, allowing for accurate and sensitive quantitative analysis. Furthermore, light emission is generally represented by steady-state(13). The TBARS is one of the oldest and most widely used methods to assess lipid oxidation in membranes. Malondialdehyde (MDA) is an oxidation product of membrane polyunsaturated acids (PUFAs) that reacts with 2-thiobarbituric acid to produce a colored complex. Other aldehydes such as alkenals and 2, 4-alkadienals also react, so it is more appropriate to use the name TBARS(14). CL and TBARS have been widely used as an indicator of ROS formation in whole cells and organs, allowing the study of a number of pathophysiological conditions related to oxidative stress.
Material and methods
Materials
Female Wistar AH/HOK were obtained from Laboratory Animal Facility, Faculty of Veterinary Science, National University of La Plata. The Institutional Committee for the Care and Use of Laboratory Animals (CICUAL) of the Faculty of Veterinary Sciences, UNLP, approved all the housing and experimentation procedures. BSA (Fraction V) was obtained from Wako Pure Chemical Industries Ltd, Japan. Alpha lipoic acid was kindly supplied by Craveri Lab S.A., Arengreen 830/Miranda 5237 C1405CYH. L (+) ascorbic acid was from Merck Laboratories. All other reagents and chemicals were of analytical grade from Sigma.
Alpha-lipoic acid solution preparation
A 200 mg of alpha lipoic acid tablet was used. It was crushed and diluted in 20 ml of distilled water, obtaining a stock solution of 10 mg/ml. 1 ml of this solution was taken and diluted in 10 ml of distilled water, thus obtaining a final concentration of 1 mg/ml. The following concentrations were used for the experiments: 50 µg/ml, 150 µg/ml and 250 µg/ml.
Animals and preparation of brain and heart microsomes
Seven-weeks-old female Wistar AH/HOK rats (n =6), weighing 120 g to 137 g were used. All rats were fed with commercial rat chow and water ad libitum. The rats were sacrificed by cervical dislocation and brains and hearts were rapidly removed, cut into small pieces and extensively washed with 0.15 M NaCl. A 30% (w/v) homogenate was prepared in a 0.25 M sucrose solution, 10 mM Tris-HCl pH 7.4 using a Potter-Elvehjem homogenizer. The homogenate was centrifuged at 1,000 x g, the pellets were discarded, and the resulting supernatant was centrifuged at 20,000 x g for 10 min. After centrifugation, 5 ml of the resulting supernatant were passed through a Sepharose 4 B column (1.6cm x 12 cm) equilibrated and diluted with 10 mM Tris-HCl buffer pH7.4 0.01% NaN3. The microsomal fraction appeared in the void volume (12-20 ml) and the cytosol was discarded. All operations were performed at 4 °C and under dim light. The quality of this microsomal preparation is compositionally similar with respect to concentrations and activities of certain microsomal enzymes to that obtained by ultracentrifugation(15).
Non-enzimatic peroxidation of rat brain and heart microsomes
The microsomes (1 mg of microsomal protein) with addition of alpha-lipoic acid solution (50, 150 and 250 µg- alpha-lipoic acid group) were incubated at 37 °C with 0.01M phosphate buffer pH 7.4, 0.4mM ascorbate, final vol. 1 ml. CL and peroxidation were initiated by adding ascorbate-Fe++ to microsomes (1 mg protein) (ascorbate-Fe++ group)(16). To carry out the experiments, three groups were established: 1) control group (only organelles), 2) ascorbate-Fe++ group (organelles + inducer) and 3) ALA group (organelles + inducer + ALA). The phosphate buffer is contaminated with enough iron to provide the necessary ferrous or ferric iron (final concentration in the incubation mixture was 2.15 μM) for peroxidation(17). Light emission from the membranes was determined over a period of 120 min, CL was recorded as cpm every 10 min, and the sum of the total CL was used to calculate cpm/mg protein. CL was measured as counts per minute on a Packard 1900 TR liquid scintillation analyzer with CL software.
Measurement of lipid peroxidation
Levels of MDA were measured using the thiobarbituric acid (TBA) fluorometric assay(18). Stock TCA-TBA-HCL reagent: 15% w/v trichloroacetic acid; 0.375% w/v thiobarbituric acid; 0.25 N hydrochloric acid. 1.0 ml of the brain or heart microsomes samples were combined with 2.0 ml of TCA-TBA-HCL and mixed well. The solution was heated for 15 min in a boiling water bath. After cooling, the flocculent precipitate was removed by centrifugation at 1000 g for 10 min. The absorbance of the sample was determined at 535 nm against a blank containing all reagents except lipid. The concentration of MDA in the sample was calculated using an extinction coefficient of 1.56 x 105 M-1 cm-1. Data were expressed as nanomoles of MDA equivalents per milligram of protein.
Protein determination
Proteins were determined by the method of Lowry(19) using BSA as a standard.
Statistical analysis
Results are expressed as mean ± S.D. of six independent determinations. Data were statistically evaluated by one way analysis of variance (ANOVA) and Tukey´s test. The statistical criterion for significance was selected at different p values, which was indicated in each case.
Results
Light emission of rat brain microsomes during peroxidation
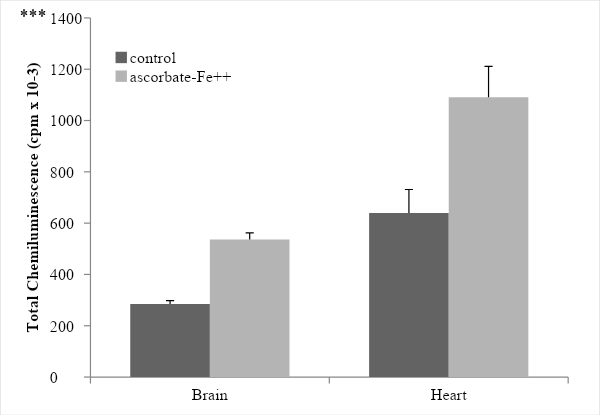
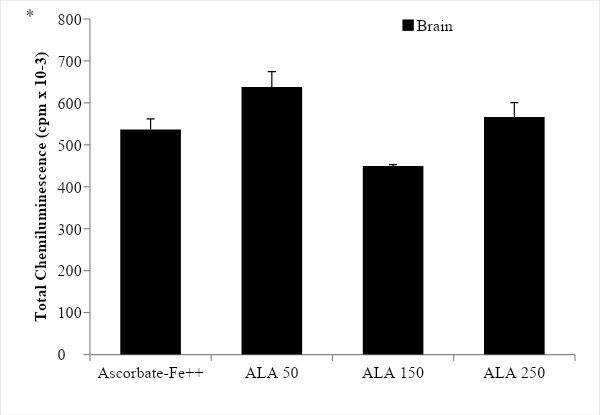
The incubation of rat brain microsomes in the presence of ascorbate-Fe++ resulted in the peroxidation of membranes as evidenced by emission of light (chemiluminescence) when the control and ascorbate-Fe++ groups were compared. The values were 285.00 ± 12.83 in the control group while 536.00 ± 25.65 in the ascorbate-Fe++ group, showing a significance value of p < 0.0005 (Figure 1). After incubation of microsomes in an ascorbate-Fe++ system at 37 °C for 120 min in the presence of increasing amounts of ALA solution (50, 150 and 250 µg- ALA group) per mg of protein, we observed that only for the 150µg/ml dose the protective effect of ALA was significant (p < 0.05). The values were 638.00 ± 36.69 with the addition of 50 µg ALA/mg protein; 449.67± 2.87 with the addition of 150 µg ALA/mg protein and finally 566.33 ± 34.24 cpm with the addition of 250 µg ALA/mg protein. The significance values were p < 0.05 with the 150 dose, while for the 50 and 250 doses no significant differences were observed (Figure 2).
Figure 1: Peroxidation ascorbate–Fe2+ of brain and heart microsomes. Chemiluminescence was determined over a 120 min period and recorded as cpm every 10 min and the sum of the total chemiluminescence was used to calculate cpm/mg protein. Results are expressed as mean ± SD of six independent experiments. Statistically significant differences between control vs. ascorbate-Fe++ group are indicated by *** p < 0.0005.
Figure 2: Light emission of rat brain microsomes during peroxidation with different concentration of ALA. The results are expressed as the mean ± S.D. of six independent experiments. Statistically significant differences between ascorbate-Fe++ vs. ALA groups are indicated by *p < 0.05.
Light emission from rat heart microsomes during peroxidation
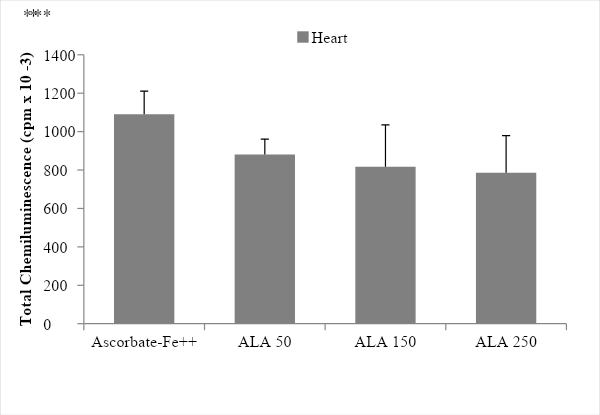
The incubation of rat heart microsomes in the presence of ascorbate-Fe++ resulted in the peroxidation of membranes as evidenced by emission of light (chemiluminescence) when the control and ascorbate-Fe++ groups were compared. The values were 639.67 ± 91.38 in the control group while 1090.67 ± 120.39 in the ascorbate-Fe++ group, showing a significance value of p < 0.0005 (Figure 1). After incubation of microsomes in an ascorbate-Fe++ system at 37 °C for 120 min in the presence of increasing amounts of ALA solution (50, 150 and 250 µg- ALA group) per mg of protein. The values were 880.33 ± 80.97 with the addition of 50 µg ALA/mg protein; 817.00 ± 218.59 with the addition of 150 µg ALA/mg protein and finally 785.33 ± 193.68 cpm with the addition of 250 µg ALA/mg protein. The significance values were p < 0.005 with the 50 dose, while for the 150 and 250 doses a highly significant value corresponding to p < 0.05 was obtained (Figure 3).
Figure 3: Light emission of rat heart microsomes during peroxidation with different concentration of ALA. The results are expressed as the mean ± S.D. of six independent experiments. Statistically significant differences between ascorbate-Fe++ vs. ALA groups are indicated by *p < 0.05, **p < 0.005.
Determination of lipid peroxidation products (thiobarbituric acid reactive substances-tbars)
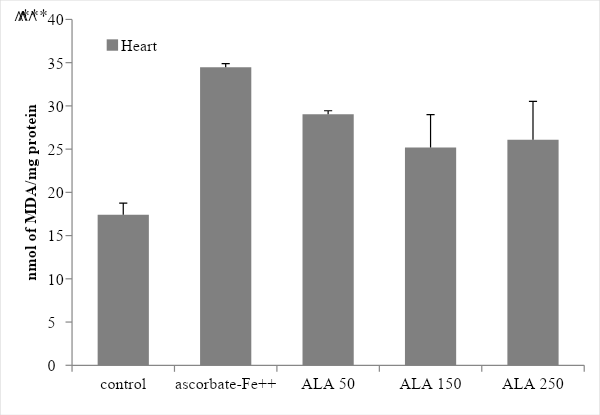
TBARS assay was used to quantify the degree of lipid peroxidation in rat brain and heart microsomes. Statistically very significant levels of MDA were found (p<0.0005) when comparing the control vs. ascorbate-Fe++ groups which indicates that in the presence of ascorbate-Fe++ (inducer) the membranes were peroxidized (Figures 4 and 5). Furthermore, when the ascorbate-Fe++ groups were compared with the ALA group in the presence of increasing concentrations of ALA (50, 150 and 250 µg AAL/mg Pt) it was observed that in brain microsomes only at the concentration 150 µg AAL/ mg Pt the concentration of MDA was significantly lower (p < 0.05) than in the ascorbate-Fe++ group (Figure 4). On the other hand, in heart microsomes it was observed that at dose 50 very significant differences were observed (p < 0.005), while at doses of 150 and 250 significant differences were observed (p < 0.05) (Figure 5).
Figure 4: Determination of Lipid Peroxidation Products (Thiobarbituric Acid Reactive Substances-TBARS) in brain microsomes: effect of ALA. Statistically significant differences between control vs. ascorbate-Fe++ group are indicated by *** p < 0.0005 and ascorbate-Fe++ vs. ALA groups are indicated by ˄p < 0.05.
Figure 5: Determination of Lipid Peroxidation Products (Thiobarbituric Acid Reactive Substances-TBARS) in heart microsomes: effect of ALA. Statistically significant differences between control vs ascorbate-Fe++ group are indicated by *** p < 0.0005 and ascorbate-Fe++ vs ALA groups are indicated by ˄˄p < 0.005, ˄p < 0.05.
Discussion
Reactive oxygen species (ROS) are produced through enzymatic and non-enzymatic pathways during normal metabolism they are part of a family of reactive species including reactive nitrogen, sulfur, carbon, selenium, electrophile and halogen (RHS) species. Supraphysiological concentrations of ROS can interact with many cellular components, including DNA, lipids, and proteins, and also generate other reactive species with potentially toxic consequences(20). Polyunsaturated fatty acids are very vulnerable to being attacked by oxygen-free radicals, which leads to lipid peroxidation(21). However, the excess or deficiency of ROS is associated with the pathogenesis of cancer, diabetes mellitus, atherosclerosis, neurodegenerative diseases, rheumatoid arthritis, ischemia/reperfusion injury, obstructive sleep apnea, and other diseases(22, 23).
Lipid peroxidation or oxidative rancidity represents a form of tissue damage that can be triggered by oxygen, singlet oxygen, hydrogen peroxide, and hydroxyl radical. Unsaturated fatty acids are essential components of cell membranes, which is why they are believed to be important for their normal functioning; however, they are vulnerable to oxidative attack initiated by oxygen free radicals( 24, 25, 26).
ALA is a powerful lipophilic antioxidant in vivo and in vitro(27). ALA differs from antioxidants in that it neutralizes free radicals both in the fatty and in the watery regions of cells, in contrast to vitamins C (water soluble) and E (fat soluble)(28). It is easily absorbed from the gastrointestinal tract, is easily transported across cell, and can cross the blood brain barrier without exhibiting any serious side effects(29). Several studies have revealed ALA protective effects in cases such as neurodegenerative diseases in which free radicals are involved(30, 31).
In this study we analyzed the in vitro antioxidant effect of ALA on non-enzymatic peroxidation in rat brain and heart microsomes in order to find an inhibitory dose that offers greater protection caused by ROS. It is important to know how biological compounds with antioxidant properties contribute to the protection of specialized membranes against deleterious effects produced by reactive oxygen species and other free radicals. With respect to previous studies carried out in our Laboratory referring to the effect of ALA on rat heart and brain mitochondria(32), we observed that when the mitochondria of both organs were incubated with different concentrations of ALA (50, 150 and 250 µg/ml), it was observed a differential protection. Rat brain mitochondria incubated with doses of 150 and 250 mg/ml of ALA were protected from the effects of non-enzymatic peroxidation (NEP), while in cardiac mitochondria, protection was only observed with the highest dose of ALA (250 µg/ml). The CL analysis indicated that rat brain mitochondria were protected more efficiently than rat heart mitochondria. But in this study, although there was a significant increase in the levels of cpm and MDA in the Fe++ ascorbate group in the microsomes of both organs, it was observed that after treatment with increasing concentrations of ALA, the analysis of CL and TBARS levels indicated that ALA acted as an antioxidant that only protects rat heart microsomes from damage by lipid peroxidation while in brain only at the 150 dose AAL acted as an antioxidant. By the other hand we observed that in brain microsomes incubated with the dose 50 µg/ml the analysis of CL and TBARS levels indicated a pro-oxidant effect of ALA.
Conclusion
Taking into account that CL and TBARS assays has been used as an indicator of ROS formation, we observed that the protective effect of ALA at the doses tested was greater in heart microsomes than in rat brain microsomes. In the latter case, it will be necessary to test new doses of ALA to demonstrate the effects on these membranes.
References
Funding
This work was supported by the Secretariat of Science and Technology, National
University of La Plata (Project 11/V276).
Credits Gavazza M, Marmunti M, Palacios: the three authors worked on conceptualization,
validation, formal analysis, research, writing, review, editing and visualization. GM and PA worked on
methodology and writing -draft original. PA worked on resources, data curation, supervision and project
administration. Conflict of interest statement The authors declare that, during the research process, there has been no type of
personal, professional or economic interest that could have influenced the judgment and/or actions of the
researchers at the time of preparing and publishing this article.