se indique lo contrario, http://www.creativecommons.org.ar/licencias.html
DOI: https://doi.org/10.19137/cienvet202325203
![]() Esta obra se publica bajo licencia Creative Commons 4.0 Internacional. (Atribución-No Comercial-Compartir Igual) a menos que
Esta obra se publica bajo licencia Creative Commons 4.0 Internacional. (Atribución-No Comercial-Compartir Igual) a menos que
se indique lo contrario, http://www.creativecommons.org.ar/licencias.html
ARTICULO DE INVESTIGACIÓN
Análisis de asociación de alelos del gen BoLA-DRB3.2 con rasgos de
producción lechera y número de células somáticas en ganado Holstein
de La Pampa
Association analysis of BoLA-DRB3.2 alleles with milk production traits and somatic cells number in La Pampa Holstein cattle
Análise de associação dos alelos do gene BoLA-DRB3.2 com características
de produção de leite e número de células somáticas em bovinos da raça
Holandesa do Pampa
Baltian LR1, Remirez P1, Peratta D1, Schmidt EE1, Palezza J1, Patrilla J1, Lema Vincens M1, Portada J1.
1Facultad de Ciencias Veterinarias. Universidad Nacional de La Pampa. Calle 5 esq. 116- General Pico, La
Pampa Argentina
Correo electrónico: lbaltian@vet.unlpam.edu.ar
Resumen: Durante muchos años el objetivo de selección en las explotaciones lecheras estuvo
focalizado en las altas producciones, ignorándose los rasgos de salud tales como la
resistencia a enfermedades. Actualmente el interés está en identificar genéticamente los
animales resistentes a desarrollar enfermedades infecciosas, por medio de genes
candidatos. El complejo principal de histocompatibilidad Bovino (BoLA) es un grupo de
genes, vinculado a la respuesta inmune. El objetivo del presente estudio fue asociar alelos
del exón 2 del gen BoLA-DRB3.2 con producción de leche y conteo de células somáticas, un
parámetro asociado a la incidencia de mastitis subclínica. Se tomaron muestras de sangre
a 157 vacas de raza Holstein y su ADN se analizó por la técnica PCR-RFLP. Se detectaron 32
alelos de los cuales seis fueron los más frecuentes (13,50 % a 6,43 %). Éstos son: BoLADRB3.2
*23, *24, *16, *25, *28 y *22. Se analizaron los conteos de células somáticas como
indicadores de enfermedad cuando los registros eran superiores a 250.000 cel/ml de leche.
Con un modelo lineal generalizado se encontró al alelo *25 asociado con bajo conteo de
células somáticas y el *23 asociado a un alto conteo de células somáticas (p= 0,03). Se
detectó una asociación entre los alelos con los litros de leche (p= 0,0132). No se encontró asociación significativa con el porcentaje de proteína y de grasa. Los alelos del BoLA DRB3.2 se evidencian como marcadores relevantes para detectar animales genéticamente resistentes a mastitis y una mayor producción lechera.
Palabras claves: Gen BoLA-DRB3.2, Resistencia a mastitis, Producción lechera, Células somáticas
Abstract For many years the selection objective on dairy farms was focused on high yields, ignoring health traits such as disease resistance. Currently the interest is focused in genetically identifying animals that are resistant to developing infectious diseases, through candidate genes. The main Bovine Histocompatibility Complex (BoLA) is a group of genes linked to the immune response. The aim of the present study was to associate alleles of exon 2 of the BoLA-DRB3.2 gene with milk production and somatic cell count as a parameter to evaluate subclinical mastitis. Blood samples were taken from 157 Holstein dairy cows; their DNA was analyzed using the PCR-RFLP technique. Thirty-two alleles were detected, six of which were the most frequent. These are: BoLA-DRB3.2 *23, *24, *16, *25, *28 and *22, with a frequency ranging from 13,50% to 6,43%. Somatic cell counts were analyzed as disease indicators when the records were above 250,000 cells/ml of milk. Using a generalized linear model, allele *25 was found to be associated with a low somatic cell count and *23 was associated with a high somatic cell count (p = 0,03). An association was detected between the alleles and liters of milk (p = 0,0132). No significant association was found with protein and fat percentages. The BoLA-DRB3.2 alleles are evidenced as relevant markers to detect animals genetically resistant to mastitis and higher milk production.
Keywords: BoLA-DRB3.2 gene, Mastitis resistance, Milk production, Somatic cells
Resumo: Por muitos anos, o objetivo da seleção em fazendas leiteiras foi focado em altos rendimentos, ignorando características de saúde, como resistência a doenças. Atualmente o interesse está em identificar geneticamente animais resistentes ao desenvolvimento de doenças infecciosas, por meio de genes candidatos. O complexo principal de histocompatibilidade bovina (bola) é um grupo de genes, ligados à resposta imune. O objetivo deste estudo foi associar os alelos do exon 2 do gene bola-DRB3.2 com a produção de leite e a contagem de células somáticas, parâmetro associado à incidência de mastite subclínica. Amostras de sangue foram coletadas de 157 vacas da raça Holandesa e seu DNA foi analisado pela técnica de PCR-RFLP. Foram detectados 32 alelos, sendo seis deles os mais frequentes (13,50% a 6,43%). São eles: bola-DRB3.2 *23, *24, *16, *25, *28 e *22. As contagens de células somáticas foram analisadas como indicadores de doença quando os registros foram superiores a 250.000 células/ml de leite. Com um modelo linear generalizado, o alelo *25 foi associado a uma baixa contagem de células somáticas e o *23 associado a uma alta contagem de células somáticas (p= 0,03). Foi detectada associação entre os alelos com os litros de leite (p= 0,0132). Não foi encontrada associação significativa com o percentual de proteína e gordura. Os alelos bola-DRB3.2 evidenciam-se como marcadores relevantes para detectar animais geneticamente resistentes à mastite e maior produção de leite.
Palavras-chave: Gene bola-DRB3.2, Resistência à mastite, Produção de leite, Células somáticas.
Durante muchos años el objetivo de selección en las explotaciones lecheras estuvo focalizado en las altas producciones por su importancia económica. Sin embargo, las producciones altas traen consecuencias en la salud y gastos en cuidados veterinarios. Los rasgos de salud generalmente tienen heredabilidad baja y cantidades limitadas de datos, lo que dificulta el potencial de mejora genética por métodos tradicionales de selección (1).
La mastitis bovina es considerada una enfermedad con elevada incidencia que afecta mundialmente la industria láctea, pues ocasiona pérdidas económicas importantes a los productores de leche debido a la disminución de la calidad y cantidad de leche producida y a un aumento en los costos por tratamiento, servicios veterinarios y pérdida de animales (2). Esta enfermedad infecciosa se caracteriza por anormalidades visibles en la ubre o en la leche. La glándula se inflama y se acumulan glóbulos blancos en la leche, que junto a las células epiteliales del tejido productor de leche, constituyen lo que se conoce como células somáticas. Las vacas con altos registros de células somáticas producen leche con menos grasa y proteína, en particular caseína. Por lo tanto, disminuye la calidad de los productos lácteos derivados. Los conteos de células somáticas (CCS) en la leche se utilizan como indicadores de la salud mamaria sobre la base de que reflejan una respuesta inmunitaria y, por lo tanto, la presencia de infección. (3) La mastitis subclínica se caracteriza por la presencia de microorganismos, un conteo elevado de células somáticas en la leche y no tener tratamiento (4). A diferencia de la mastitis clínica, en este caso no existen alteraciones obvias en la ubre (5). No obstante, sí ocurren cambios en la composición de la leche a causa de las bacterias y componentes inflamatorios (6).
La necesidad de una mayor eficiencia en el proceso industrial de la leche y la creciente demanda del mercado por productos de mayor calidad traen como consecuencia un incremento en las exigencias de los estándares de la materia prima. Habitualmente se hace referencia a un precio pagado por litro de leche producido y en realidad el mismo surge de la conjunción de diferentes parámetros, uno de los cuales está referido a la calidad sanitaria de la leche, valorado a través del registro de células somáticas presentes. Si la calidad de la leche no es buena hay pérdidas económicas (7) ya que se afecta el rendimiento del queso y la vida útil de todos los productos lácteos (8, 9 ).
Es por ésta razón que desde hace varios años existe interés, no sólo en seleccionar a favor de una alta producción y calidad lechera (litros de leche, cantidad y porcentaje de grasa y proteínas), sino también en seleccionar a favor de un bajo CCS.
Una posible solución es identificar loci de rasgos cuantitativos que controlan rasgos sanitarios, como marcadores genéticos y facilitar la selección para mejorar sanidad y aptitud en las vacas lecheras y así seleccionar animales resistentes a enfermedades infecciosas. La variabilidad genética de la resistencia a la mastitis está detectada en el ganado lechero. Muchos estudios se centraron en la variación poligénica del rasgo, al estimar la heredabilidad y la correlación genética entre los rasgos fenotípicos relacionados con la mastitis, tales como los recuentos de células somáticas y los casos clínicos (10).
La variación en el resultado de la infección depende en gran medida de las diferencias en la constitución genética de los individuos. Los factores que pueden influenciar el resultado de la infección son varios y se clasifican en: variables ambientales; variables propias del patógeno y factores del hospedador (11). Generalmente, las variables ambientales en la resistencia/susceptibilidad a enfermedades infecciosas tienen un efecto considerable, por lo que estos caracteres presentan una mediana a baja heredabilidad.
Aquellas variables derivadas del patógeno, implican factores de virulencia que pueden incrementar o atenuar la habilidad de colonización del organismo, o aumentar la patogenicidad a través de la expresión de una toxina. Finalmente, los factores atribuidos al hospedador implican principalmente la dotación genética del organismo dado que en toda población existen diferencias individuales en la resistencia/susceptibilidad a enfermedades infecciosas (11).
Los loci del Complejo Mayor de Histocompatibilidad (MHC) constituyen los principales genes candidatos para el estudio de asociación entre marcadores genéticos y resistencia/susceptibilidad a enfermedades infecciosas.
Cada uno de sus genes a su vez tiene muchos alelos por lo que es también polimórfico. Este polimorfismo tiene un profundo efecto en el reconocimiento de los antígenos por las células T y la combinación de poligenia y polimorfismo extiende ampliamente el rango de péptidos que pueden ser presentados a las células T por un individuo (12).
En bovinos el MHC se denomina Bovine Leukocyte Antigen (BoLA) y ha sido mapeado en el autosoma bovino 23 (BTA 23) dentro de una extensión de 4.000 Kb que contiene más de 154 genes estrechamente ligados (13). Está estructurado en moléculas de Clase I, II y III. Las analizadas en este estudio son las moléculas de clase II, compuestas de un locus DRA y al menos 3 loci DRB. Entre estos loci, el gen DRB3 es altamente polimórfico y en particular el exón 2 (BoLA-DRB3.2) de dicho gen codifica muchos rasgos relacionados con la inmunidad a enfermedades (14).
En vacas lecheras los polimorfismos de estas moléculas se han asociado con resistencia/susceptibilidad a enfermedades infecciosas tales como mastitis clínica, leucosis, herpes virus 1 (Bo-HV1) y neosporosis entre otras (15-20). Sin embargo, es poco el avance de las investigaciones para conocer si los loci específicos del BoLA de clase II están individualmente asociados con bajos y altos CCS, como medida de resistencia/susceptibilidad, o si otros genes ligados son responsables de dicha asociación. Por lo tanto, el objetivo fue analizar los polimorfismos en el locus de Clase II BoLA-DRB3.2 y asociarla con rasgos de producción lechera y CCS en ganado Holstein de La Pampa. Los objetivos específicos fueron: a) Caracterizar los polimorfismos presentes en el locus de Clase II BoLA-DRB3.2 en la población en estudio. b) Analizar la asociación entre el CCS y los alelos del locus de Clase II BoLA-DRB3.2. c) Asociar datos de producción de litros de leche, porcentaje de grasa y proteína y los alelos del locus de Clase II BoLA-DRB3.2.
Los animales estudiados pertenecen a un tambo de la zona aledaña a General Pico, departamento Maracó, La Pampa. Se tomaron muestras de 5 ml de sangre a 157 vacas lecheras de la raza Holstein. Dichas muestras se conservaron con EDTA a -20 °C hasta su utilización.
De los animales muestreados se obtuvieron registros promedios mensuales de conteo de células somáticas y registros de producción tales como litros de leche, porcentaje de grasa y porcentaje de proteína durante un periodo de cuatro años.
Diseño experimental
El registro de CCS se transformó a una variable dicotómica de 0 y 1. Representado 0 en el caso de aquellos individuos que poseían menos de 250.000 células somáticas por mililitro leche y por 1 a aquellos que superaban dicho valor (CCS ≥ 250.000 cel/ml), (NMC Global Milk Quality http://www.nmconline.org). Cada registro de producción de leche, porcentaje de grasa y porcentaje de proteína es el promedio mensual de ese rasgo productivo. Estos datos se relevaron mensualmente durante un lapso de tiempo de 4 años y el total de lecturas de cada animal varió entre 10 y 30.
Extracción de ADN
El ADN genómico se extrajo a partir de las muestras de sangre entera mediante el kit comercial de HIGHWAY (ADN Puri Prep-S Kit). El ADN genómico extraído se analizó en geles de agarosa. Se tomaron 3 ul de cada muestra y se mezclaron con 3 ul de colorante. La electroforesis se realizó a 100 voltios durante 30 minutos. Los geles se sumergieron en un baño con bromuro de etidio durante 10 minutos y se observaron en transiluminador de emisión de luz ultravioleta (UV).
Tipificación del exón 2 del gen BoLA-DRB3.2 mediante PCR-RFLP
La caracterización de los alelos del locus BoLA-DRB3 fue llevada a cabo por el método de PCR anidado-RFLP descrito por van Eijk et al. (21). Dicha técnica consiste en la amplificación de una secuencia de ADN de 284 pb del exón 2 del gen BoLA-DRB3 y la posterior digestión de los productos de PCR con las enzimas de restricción Rsa I, Bsty I y Hae III. La clasificación de los alelos se realizó teniendo en cuenta las combinaciones de los patrones de restricción obtenidos en forma independiente para las enzimas antes mencionadas.
Reacción en cadena de la polimerasa (PCR)
En la primera ronda de amplificación se utilizaron los cebadores HL030 (5´-ATCCTCCTCTCTGCAGCACATTTCC-3´) y HL031 (5´-TTTAAATTCGCGCTCACC TCGCC GCT-3´). El programa utilizado en la reacción de PCR consistió en 4 minutos a 94° C de desnaturalización inicial, 9 ciclos de 1 minuto a 94°C, 2 minutos a 60° C y 1 minuto a 72°C, seguidos por 4 minutos de polimerización final a 72ºC.En la segunda etapa se usaron los cebadores HL030 y HL032 (5´- TCGCCGCTGCACAGTGAAACTCT -3´) (Lewin et al. 1992). El programa consistió en: una etapa de desnaturalización de 1 minuto a 94°C seguida de 19 ciclos de 1 minuto a 94°C, 30 segundos a 65°C y 1 minuto a 72° C, seguidos por 4 minutos de polimerización final a 72ºC.
Polimorfismos de la longitud de los fragmentos de restricción (RFLP)
Alícuotas del producto de amplificación (12 μl) se digirieron por separado con las enzimas de restricción Rsa I, Hae III y Bsty I (5 U) durante 3 horas, de acuerdo con las indicaciones del proveedor (Promega, Madison, WI; New England Biolabs, Beverly, MA, USA). Las temperaturas de digestión fueron de 37°C para las dos primeras enzimas y de 60°C para la tercera. Los fragmentos de restricción se resolvieron en geles de poliacrilamida 8% que se utilizaron para separar las bandas de los productos de la digestión enzimática (PCR-RFLP). Los geles se tiñeron con nitrato de plata.
Análisis estadístico
Se utilizó un modelo lineal generalizado para el análisis de asociación entre la variable respuesta como rasgo productivo y los alelos. Se tuvieron en cuenta los efectos fijos como la estación del año y el año de registro y alelos BoLA, además, se consideraron los efectos aleatorios de cada animal como son la producción, el porcentaje de grasa y el porcentaje de proteínas. La variable respuesta será uno de los rasgos productivos; la producción de leche, el porcentaje de grasa y el porcentaje de proteína. Este rasgo productivo estará asociado a un promedio general de dicho rasgo, al efecto fijo de la estación, al efecto fijo del año, al efecto aleatorio animal sobre el rasgo analizado con media 0 y varianza igual a la covarianza entre mediciones repetidas dentro de cada animal y a un error con media 0 y varianza σ2.
Para analizar el CCS se transformó la variable a una variable dicotómica, tomando el valor 1 para el caso en que el CCS> 250.000cel/ml (mastitis subclínica) y tomando el valor 0 para el caso en que el CCS< 250.000cel/ml. Esta variable dicotómica se analizó mediante un modelo de regresión logística cuya función de enlace relaciona el valor esperado con el predictor lineal (ƞi) y viene expresada mediante la siguiente ecuación log(πi / (1 - πi)) = ƞi. El valor del predictor lineal es la probabilidad de ocurrencia de la mastitis subclínica asociado a una media general y los demás efectos que fueron considerados en el modelo anterior de los rasgos productivos. Se menciona que el modelo se ajustó en función del criterio de Akaike (1974).
En ambos modelos se utilizó el software InfoStat el cual incluye librerías del software R para la resolución de los problemas.
En la población en estudio se caracterizaron los polimorfismos presentes en el exón 2 del gen BoLA-DRB3 por PCR-RFLP y se identificaron un total de 32 alelos distintos, en un muestreo de 157 vacas. Los alelos de mayor frecuencia hallados son el alelo DRB3.*23,*24, *16, *25, *28 y *22, todos tuvieron una frecuencia absoluta mayor o igual a 20, tal como se muestra en la Tabla N° 1.
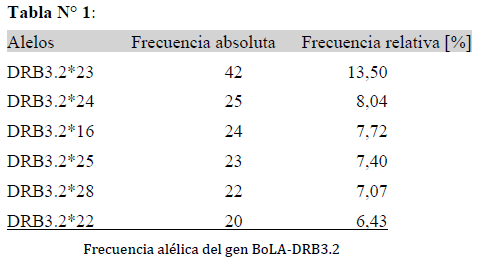
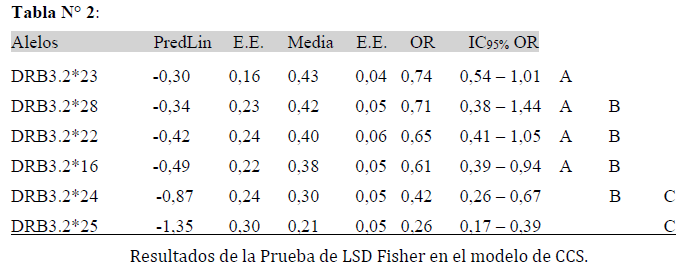
Los resultados de la asociación entre el conteo de células somáticas y alelos indicaron efectos significativos en la asociación (p= 0,03). El modelo planteado permitió hallar las probabilidades que resultan de aplicar la transformación inversa de la función enlace del modelo de regresión logística. A su vez, los Odds Ratio se calculan por OR=ePredLin. La prueba LSD Fisher evidenció diferencias entre los alelos.
La Tabla N° 2 muestra estos resultados y además el correspondiente intervalo de confianza del 95% para los Odds Ratio. Estas diferencias en el CCS muestra a los alelos BoLA-DRB3.2*23, *28, *22 y *16 identificados con la letra A con incidencia estimada de (altos conteos de células somáticas) entre el 38% y el 43% de probabilidad de tener mastitis subclínica y cuyos OR varían entre 0,61 y 0,74. Los alelos identificados con la letra B son BoLA-DRB3.2*28, *22, *16 y *24, los tres primeros también se encuentran identificados con A, con incidencias estimadas entre el 30% y el 42% y sus OR entre 0,42 y 0,71. Y por último identificados con la letra C los alelos BoLA-DRB3.2*24 y *25 con incidencia del 30% y 21% son los alelos que poseen la menor probabilidad de poseer mastitis subclínica y cuyos OR son 0,42 y 0,26.
De los rasgos productivos analizados y su error estándar se obtuvieron estos valores promedios, para la producción 19,49 litros (0,16 litros), para el porcentaje de grasa 4,03% (0,02%) y para el porcentaje de proteína 3,54% (0,10%).
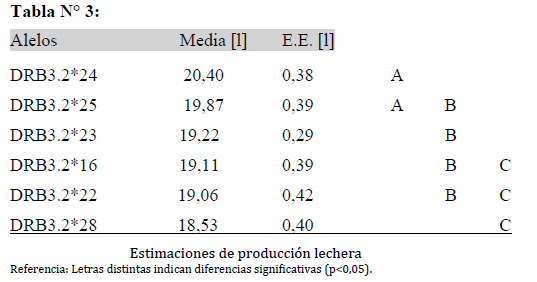
Del modelo planteado se observó una asociación entre los alelos con la producción de leche (p= 0,0132) y no se encontró asociación significativa con el porcentaje de proteína y de grasa.
La Tabla N° 3 muestra las estimaciones de producción promedio de leche y su correspondiente error estándar para cada alelo analizado. Se diferencian los niveles de producción identificados con la letra A, B y C. Mostrando que los alelos BoLA-DRB3.2*24 y BoLA-DRB3.2*25 identificados con la letra A tienen el mayor nivel de producción promedio de leche, mientras que los alelos BoLA-DRB3.2*16, BoLA-DRB3.2*22 y BoLA-DRB3.2*28 identificados con la letra C tienen el menor nivel de producción promedio de leche.
En el presente trabajo se caracterizaron los polimorfismos presentes en el exón 2 del gen BoLA-DRB3 en una población de bovinos de raza Holstein de la provincia de La Pampa y su asociación con CCS y rasgos de producción de leche. El exón 2 es altamente polimórfico y es reconocido por su rol en la resistencia/susceptibilidad a enfermedades infecciosas y autoinmunes. Por este motivo han sido llevados a cabo numerosos estudios en diferentes razas bovinas, tanto criollas como comerciales, utilizando diferentes metodologías (14, 23-30, 31, 32).
En nuestro estudio los resultados obtenidos también evidenciaron un elevado polimorfismo en dicho gen, detectando un total de 32 alelos. Todas las variantes encontradas habían sido reportadas previamente. Dicha coincidencia de diversidad alélica de las diferentes razas bovinas sigue manifestándose a pesar de la alta presión de selección sufrida por la raza Holstein y el alto nivel de consanguinidad(29, 22, 26).
Los alelos de mayor frecuencia (n≥ 20) hallados son el alelo BoLA-DRB3.*23, *24, *16, *25, *28 y *22, en coincidencia parcial con lo reportado por Pokorska et al. (28) quienes detectaron que el alelo BoLA-DRB3 *24, *22 y *16 eran los más frecuentes en su estudio. Estos resultados podrían ser consecuencia del alto nivel de homogeneidad exhibido por esta raza debido al uso de la misma genética a nivel global (29).
Asociación entre los alelos del locus BoLA-DRB3 y CCS
Numerosos estudios han analizado la asociación entre los alelos del gen BoLA-DRB3 y la susceptibilidad/resistencia a varias enfermedades infecciosas, focalizándose principalmente en mastitis, leucosis bovina y dermatofilosis, entre otras (14, 16-19, 23, 25, 30).
De acuerdo a los resultados obtenidos en el presente trabajo se detectó que el alelo BoLA-DRB3.2*23 estaba asociado con alto conteo de células somáticas CCS (p=0,03), siendo considerados dentro de este grupo aquellos animales que habían desarrollado mastitis o bien poseían ≥250.000 cel/ml. Este resultado es coincidente con lo reportado por Shariff et al. (30) Rupp et al. (32) y Oprzadek et al. (33). En tanto, Yoshida et al. (22), Baltian et al. (16), Zambrano et al. (35) y Dietz et al. (14) en oposición a lo mencionado, lo encontraron asociado con resistencia a mastitis.
En este estudio el alelo DRB3.2*28, que poseía una frecuencia considerable para estudiarlo, se lo encontró en el grupo de mayor CCS y de menor producción lechera. En tanto que Suprovych et al. (26) y Oprzadek et al. (33) asociaron al alelo DRB3.2*28, a un bajo conteo de células somáticas.. En nuestro estudio el alelo DRB3.2*16 pertenece al mismo grupo que el alelo *23 (Tabla N°2) se encontró asociado con mayor CCS al igual que Baltian et al. (16), Dietz et al. (14) y Yoshida et al. (22, 37) quienes reportaron que el BoLA DRB3.2*16 se asociaba con elevado CCS y determinaron que el mismo actuaba como factor de riesgo para el incremento agudo y crónico en el CCS en vacas Holstein de segundo parto. Por lo contrario Pokorska et al. (28), Shariff et al. (30) y Zambrano et al. (35) lo asociaron a un bajo riesgo de mastitis clínica. Rupp et al. (32) no encontraron asociaciones entre el alelo DRB3.2*16 y CCS.
El alelo BoLA-DRB3.2*24 en este trabajo se asoció con disminución del CCS en concordancia con lo propuesto por Rupp et al. (32) Kulberg et al. (36). Por otro lado, Starkenburg et al. (1) Shariff et al. (30) y Yoshida et al. (22) informaron asociación entre el alelo BoLA-DRB3*24 con la enfermedad. En este estudio el alelo DRB3.2*25 mostró una mayor asociación con bajos conteos de células somáticas (p= 0,03), tal cómo se muestra en la Tabla N°2.
Asociación entre los alelos del locus BoLA-DRB3 y rasgos productivos
Los alelos del BoLA también han sido vinculados con caracteres de producción lechera tales como producción de leche, proteína y grasa en leche (15, 35, 38).
En cuanto a la asociación con rasgos productivos en este estudio se encontró asociación del alelo*25 con una mayor producción de leche. Si bien no se encontró un dato similar en la bibliografía, esta asociación resulta un hallazgo importante. El alelo DRB3.2 *16 está en el grupo de menor producción de leche (p= 0,0132) (Tabla N°3) al igual que lo reportado por Rupp et al. (32). En este estudio, no se encontró asociación significativa con el porcentaje de proteína en la producción de leche. En tanto que Pashmi et al. (39) encontraron que los alelos BoLA-DRB3.2*22 y DRB3.2*24, están asociados a un alto porcentaje de proteína, siendo lo contrario para Shariff et al. (30). Con respecto al rasgo productivo porcentaje de grasa en la leche, en este trabajo no se encontró asociación significativa, en tanto que Starkenburg et al. (1) y Pashmi et al. (39) encontraron al alelo DRB3.2*22 asociado a un aumento de grasa durante la primera lactancia. El DRB3.2*11 también fue asociado con producción de grasa por Pashmi et al. (39).
Los resultados en general concuerdan con las frecuencias alélicas de las poblaciones de Holstein estudiadas por otros autores. El alto polimorfismo en el locus estudiado puede tener un efecto significativo en la resistencia a la mastitis.
Los animales portadores del alelo BoLA-DRB3.2*25 se presentan como los más resistentes a la mastitis y con una mayor producción de litros de leche, lo cual es un resultado interesante para nuestro rodeo.
Los estudios de asociación entre los alelos del gen BoLA-DRB3, mastitis y rasgos de producción, se constituyen como una efectiva herramienta para el control de la enfermedad y para seleccionar animales de mayor producción sugiriendo su inclusión en los programas de selección genética.
1. Starkenburg RJ, Hansen LB, Kehrli ME Jr, Chester-Jones H. Frequencies and effects of alternative DRB3.2 alleles of bovine lymphocyte antigen for Holsteins in milk selection and control lines. J Dairy Sci [Internet]. diciembre de 1997;80(12):3411-9. Disponible en: http://dx.doi.org/10.3168/jds.S0022-0302(97)76316-1
2. Fernández Bolaños OF, Trujillo Graffe JE, Peña Cabrera JJ, Cerquera Gallego J, Granja Salcedo Y. Mastitis bovina: generalidades y métodos de diagnóstico. Revista electrónica de Veterinaria [Internet]. 2012;13(11):1-20. Disponible en: https://www.produccion-animal.com.ar/sanidad_intoxicaciones_metabolicos/infecciosas/bovinos_leche/78-mastitis.pdf
3. Bradley A., Green M. 2005. Use and interpretation of somatic cell count data in dairy cows. In practice. 27: 310-315. Disponible:https://bvajournals.onlinelibrary.wiley.com/doi/abs/10.1136/inpract.27.6.310
4. Quevedo W. Recuento de celulas somaticas (rsc), como indicador en la resistencia de la mastitis bovina. Revista Ciencia, Tecnología e Innovación [Internet]. 2018; Disponible en: http://www.scielo.org.bo/scielo.php?pid=S2225-87872018000100005&script=sci_arttext
5. Kerr DE, Wellnitz O. Mammary expression of new genes to combat mastitis. J Anim Sci [Internet]. 2003;81 Suppl 3(15 Suppl 3):38-47. Disponible en: http://dx.doi.org/10.2527/2003.81suppl_338x
6. Blum SE, Heller ED, Leitner G. Long term effects of Escherichia coli mastitis. Vet J [Internet]. julio de 2014;201(1):72-7. Disponible en: http://dx.doi.org/10.1016/j.tvjl.2014.04.008
7. Guimarães JLB, Brito MAV, Lange CC, Silva MR, Ribeiro JB, Mendonça LC, et al. Estimate of the economic impact of mastitis: A case study in a Holstein dairy herd under tropical conditions [Internet]. Vol. 142, Preventive Veterinary Medicine. 2017. p. 46-50. Disponible en: http://dx.doi.org/10.1016/j.prevetmed.2017.04.011
8. Bedolla CC, de León MP. Pérdidas económicas ocasionadas por la mastitis bovina en la industria lechera. REDVET Revista electrónica de Veterinaria [Internet]. 2008;9(4):1-26. Disponible en: https://www.redalyc.org/pdf/636/63611952010.pdf
9. Tirado PO, Rojas JRA, Espinoza MM, Falcón VV, Salas RG, Andrade RM. Mastitis bovina y su repercusión en la calidad de la leche. Revista Electrónica de Veterinaria [Internet]. 2017;18(11):1-16. Disponible en: https://www.redalyc.org/pdf/636/63653574004.pdf
10. Rupp R, Boichard D. Genetics of resistance to mastitis in dairy cattle. Vet Res [Internet]. Sep-Oct de 2003;34(5):671-88. Disponible en: http://dx.doi.org/10.1051/vetres:2003020
11. Díaz S, Ripoli MV, Peral García P, Giovambattista G. Marcadores genéticos para resistencia y susceptibilidad a enfermedades infecciosas en animales domésticos. Analecta Veterinaria [Internet]. 2005;25. Disponible en: http://sedici.unlp.edu.ar/handle/10915/11177
12. Brown JH, Jardetzky TS, Gorga JC, Stern LJ, Urban RG, Strominger JL, et al. Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1 [Internet]. Vol. 364, Nature. 1993. p. 33-9. Disponible en: http://dx.doi.org/10.1038/364033a0
13. Bovine Genome Sequencing and Analysis Consortium, Elsik CG, Tellam RL, Worley KC, Gibbs RA, Muzny DM, et al. The genome sequence of taurine cattle: a window to ruminant biology and evolution. Science [Internet]. 24 de abril de 2009;324(5926):522-8. Disponible en: http://dx.doi.org/10.1126/science.1169588
14. Dietz AB, Detilleux JC, Freeman A E, Kelley D H, Stabel JR, Kehrli M E. Genetic association of bovina lymphocyte antigen DRB3 alleles with immunological traits of Holstein cattle. J Dairy Sci. 1997;(80):400-5. Disponible en: https://doi.org/10.3168/jds.S0022-0302(97)75950-2
15. Takeshima SN, Aida Y. Structure, function and disease susceptibility of the bovine major histocompatibility complex. Anim Sci J [Internet]. abril de 2006;77(2):138-50. Disponible en: http://doi.wiley.com/10.1111/j.1740-0929.2006.00332.x
16. Baltian LR, Ripoli MV, Sanfilippo S, Takeshima SN, Aida Y, Giovambattista G. Association between BoLA-DRB3 and somatic cell count in Holstein cattle from Argentina. Mol Biol Rep [Internet]. julio de 2012;39(7):7215-20. Disponible en: http://dx.doi.org/10.1007/s11033-012-1526-y
17. Baltian LR, Follmer AV, Peratta DL, Schmidt EE, Severini RA, Borrego C, et al. Polimorfismos del exón 2 del gen BoLA-DRB3 asociados con resistencia/susceptibilidad a leucosis en ganado Holstein de La Pampa. Ciencia Veterinaria [Internet]. 2016;18. Disponible en: http://sedici.unlp.edu.ar/handle/10915/105572
18. Polat M, Takeshima SN, Aida Y. Epidemiology and genetic diversity of bovine leukemia virus. Virol J [Internet]. 2 de noviembre de 2017;14(1):209. Disponible en: http://dx.doi.org/10.1186/s12985-017-0876-4
19. Morales JPA, López-Herrera A, Zuluaga JE. Association of BoLA DRB3 gene polymorphisms with BoHV-1 infection and zootechnical traits. Open Vet J [Internet]. octubre de 2020;10(3):331-9. Disponible en: http://dx.doi.org/10.4314/ovj.v10i3.12
20. Schwab AE, Geary TG, Baillargeon P, Schwab AJ, Fecteau G. Association of BoLA DRB3 and DQA1 alleles with susceptibility to Neospora caninum and reproductive outcome in Quebec Holstein cattle. Vet Parasitol [Internet]. 28 de octubre de 2009;165(1):136-40. Disponible en: https://doi.org/10.1016/j.vetpar.2009.07.004
21. van Eijk MJ, Stewart-Haynes JA, Lewin HA. Extensive polymorphism of the BoLA-DRB3 gene distinguished by PCR-RFLP. Anim Genet [Internet]. 1992;23(6):483-96. Disponible en: http://doi.wiley.com/10.1111/j.1365-2052.1992.tb00168.x
22. Yoshida T, Mukoyama H, Furuta H, Kondo Y, Takeshima SN, Aida Y, et al. Association of the amino acid motifs of BoLA-DRB3 alleles with mastitis pathogens in Japanese Holstein cows. Anim Sci J [Internet]. octubre de 2009;80(5):510-9. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1740-0929.2009.00664.x
23. Baltian LR, Rípoli MV, Giovambattista G. Determinación de los motivos aminoacídicos presentes en los sitios de unión a los antígenos de los alelos del gen BoLA-DRB3 en una población Holstein de La Pampa y su asociación con mastitis. Ciencia veterinaria. 2014; 16(1):9-27.Disponible en:https://cerac.unlpam.edu.ar/index.php/veterinaria/article/download/1719/1701
24. Gilliespie BE, Jayarao BM, Dowlen HH, Oliver SP. Analysis and frequency of bovine lymphocyte antigen DRB3.2 alleles in Jersey cows. J Dairy Sci [Internet]. septiembre de 1999;82(9):2049-53. Disponible en: http://dx.doi.org/10.3168/jds.S0022-0302(99)75443-3
25. Park YH, Joo YS, Park JY, Moon JS, Kim SH, Kwon NH, et al. Characterization of lymphocyte subpopulations and major histocompatibility complex haplotypes of mastitis-resistant and susceptible cows. J Vet Sci [Internet]. marzo de 2004;5(1):29-39. https://doi.org/10.4142/jvs.2004.5.1.29
26. Suprovych TM, Vishchur OI, Suprovych MP, Chepurna VA. Relationship between alleles of gene BoLA-DRB3 and somatic cells amount in milk of Ukrainian black-and-white dairy breed. Anim Biol Leiden Neth [Internet]. 2019;21(4):75-83. Disponible en: http://aminbiol.com.ua/20194pdf/11.pdf
27. Miyasaka T, Takeshima SN, Matsumoto Y, Kobayashi N, Matsuhashi T, Miyazaki Y, et al. The diversity of bovine MHC class II DRB3 and DQA1 alleles in different herds of Japanese Black and Holstein cattle in Japan. Gene [Internet]. 1 de febrero de 2011;472(1-2):42-9. Disponible en: http://dx.doi.org/10.1016/j.gene.2010.10.007
28. Pokorska J, Kułaj D, Dusza M, Ochrem A, Makulska J. The influence of BoLA-DRB3 alleles on incidence of clinical mastitis, cystic ovary disease and milk traits in Holstein Friesian cattle. Mol Biol Rep [Internet]. octubre de 2018;45(5):917-23. Disponible en: http://dx.doi.org/10.1007/s11033-018-4238-0
29. Baltian L, Ripoli MV, Aida Y, Takeshima SN, Giovambattista G. Estimación de las frecuencias alélicas del gen BoLA-DRB3 en una población de ganado Holstein de La Pampa mediante secuenciación directa. 2011 [citado 16 de febrero de 2023]; Disponible en: https://repo.unlpam.edu.ar/handle/unlpam/4358
30. Sharif S, Mallard BA, Wilkie BN, Sargeant JM, Scott HM, Dekkers JC, et al. Associations of the bovine major histocompatibility complex DRB3 (BoLA-DRB3) alleles with occurrence of disease and milk somatic cell score in Canadian dairy cattle. Anim Genet [Internet]. junio de 1998;29(3):185-93. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1365-2052.1998.00318.x
31. Giovambattista G, Golijow DC, Dulout FN y Lojo MM. Gene frequencies of DRB3.2 locus of Argentine Creole cattle. Anim Genet. 1996; 27:55-56. Disponible en: https://doi.org/10.1111/j.1365-2052.1996.tb01178.x
32. Rupp R, Hernandez A, Mallard BA. Association of bovine leukocyte antigen (BoLA) DRB3.2 with immune response, mastitis, and production and type traits in Canadian Holsteins. J Dairy Sci [Internet]. febrero de 2007;90(2):1029-38. Disponible en: http://dx.doi.org/10.3168/jds.S0022-0302(07)71589-8
33. Oprzadek JM, Brzozowska AM, Urtnowski P, Rutkowska K, Lukaszewicz M. Association of BoLA-DRB3 genotype with somatic cell count in milk of Polish Holstein cattle. Rev Bras Zootec [Internet]. 25 de junio de 2018 [citado 16 de febrero de 2023];47(0). Disponible en: https://www.scielo.br/j/rbz/a/8qhn97wwsV3NyH3zSrqybLB/abstract/?lang=en
34. Yoshida T, Furuta H, Kondo Y, Mukoyama H. Association of BoLA-DRB3 alleles with mastitis resistance and susceptibility in Japanese Holstein cows. Anim Sci J [Internet]. mayo de 2012;83(5):359-66. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1740-0929.2011.00972.x
35. Zambrano JC, Echeverri JZ, López-Herrera A. Alelos del gen BoLA DRB3.2 están asociados con mastitis en vacas lechera. Rev Colom Cienc Pecu. 2011;24(2):145-56. Disponible en: http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-06902011000200006
36. Kulberg S, Heringstad B, Guttersrud OA, Olsaker I. Study on the association of BoLA-DRB3.2 alleles with clinical mastitis in Norwegian Red cows. J Anim Breed Genet [Internet]. agosto de 2007;124(4):201-7. Disponible en:https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1439-0388.2007.00662.x
37. Yoshida T, Mukoyama H, Furuta H, Holmes CW, Kosugiyama M, Tomogane H. Allelic frequency of PCR-RFLP type of the BoLA-DRB3 in Japanese Holstein herds and the relation to mastitis. Anim Sci J [Internet]. agosto de 2008;79(4):409-16. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1740-0929.2008.00545.x
38. Machado MA, Nascimento CS, Martinez ML, Silva M, Campos AL, Teodoro RL, et al. Associação do loco BoLA-DRB3. 2 com produção de leite em bovinos da raça Gir. Arq Bras Med Vet Zootec [Internet]. 2005;57:380-9. Disponible en: https://www.scielo.br/j/abmvz/a/XgXf9fqpgP8ysLPR5wRdv5B/citation/?lang=pt
39. Pashmi M, Qanbari S, Ghorashi SA, Sharifi AR, Simianer H. Analysis of relationship between bovine lymphocyte antigen DRB3.2 alleles, somatic cell count and milk traits in Iranian Holstein population. J Anim Breed Genet [Internet]. agosto de 2009;126(4):296-303. Disponible en: http://dx.doi.org/10.1111/j.1439-0388.2008.00783.x
Contribuciones de autor: Todos los autores contribuyeron a la concepción y el diseño del estudio
Conflictos de intereses: Los autores declaran por escrito, no tener vínculo o compromiso que condicione lo expresado en el artículo de su autoría y que pueda ser entendido como conflicto de intereses.
Fecha de recepción: 24 de Febrero de 2023
Fecha de aceptación: 04 de Mayo de 2023